Did extinct these animals have cheeks? Paleontologists work from fossils of dinosaurs, so how can they know? An anatomist explains.
By Ali Nabavizadeh (@Vert_Anatomist)
An animal’s anatomy can tell us a lot about how it lived, including how it moved, how it ate, how it breathed, and just about any other physiological process involving morphology. In studying modern vertebrates, performing dissections is essential for understanding anatomical similarities and differences between various species and what these comparisons can tell us about their evolutionary relationships. Dissection can reveal how an animal’s internal structures worked together in life, such as how muscles and bones connected and worked. Comparative anatomists use dissection to study all types of vertebrates, but the vertebrate paleontologists among them are at a bit of disadvantage. Namely, the bones of their extinct study subjects are all that are left for a vertebrate paleontologist to work with when studying extinct dinosaurs. This makes questions such as “Did plant-eating dinosaurs have cheeks?” especially hard to answer.
The problem with fossils
Soft tissue rarely is fossilized because of how quickly it breaks down after death. This process usually leaves just the animal’s fossilized bones for vertebrate paleontologists to study, save a few exceptions. Because of this limitation, many vertebrate paleontologists use techniques such as the extant phylogenetic bracketing (EPB) to reconstruct soft tissues (Witmer, 1995). The EPB uses what is known of the anatomy of related living animal species to assess the likelihood of the existence and placement of certain soft tissue structures in the extinct animal. In addition to the EPB, there are also functional adaptations that need to be taken into account that may be outside of what is known of living relative species. This conceptual fusion of evolutionary relationships and functional adaptations is what vertebrate paleontologists use to reconstruct soft tissue anatomy to deduce functional morphology. Incidentally, these techniques have been instrumental as we investigate form and function in one of the most mysterious and popular groups of extinct vertebrates—the dinosaurs (e.g., Holliday, 2009).

The horned ceratopsian dinosaur Chasmosaurus (left) at the Academy of Natural Science in Philadelphia, Pennsylvania; the duck-billed hadrosaur Edmontosaurus (right) at the Denver Museum of Science and Nature.
My research in particular investigates the functional anatomy and evolution of muscles around the heads of the largely plant-eating, or herbivorous, group of dinosaurs known as ornithischians (Fig. 1). The ornithischian clade consists of the horned ceratopsians (e.g., Triceratops), the plated stegosaurs (e.g., Stegosaurus), the tank-like armored ankylosaurs (e.g., Ankylosaurus), the dome-headed pachycephalosaurs, the so-called “duck-billed” hadrosaurs, and their relatives. In my opinion, what makes these animals so fascinating is the incredible diversity of feeding adaptations they had acquired. Just as large plant-eating mammals do today, these dinosaurs used fine-tuned, specialized heads and teeth to eat diverse plant matter all around the world. This study of mine (Nabavizadeh, 2018) looks at reassessing the head muscles in ornithischian dinosaurs to investigate, not only their feeding function, but whether or not they needed “cheeks” to facilitate their feeding as mammals do.
So, did these dinosaurs have cheeks?
For over a hundred years, there has been an ongoing debate about the presence or absence of so-called “cheeks” in ornithischian dinosaurs (see Galton [1973] for a summary). These “cheeks” were thought to be present because of an inset tooth row in these animals allowing room for such a structure on the outside of the mouth. Although at its surface it might seem insignificant, it has actually been a point of contention among paleontologists as we try to figure out how ornithischian “cheeks” would have looked. The original idea was that the muscle fibers of the “cheeks” were oriented straight up and down between the upper and lower jaws along prominent outer ridges, acting like the buccinators (i.e., cheek muscles) in mammals to help keep food in the mouth.
The problem was that this muscle was totally made up and is only known in mammals (not in reptiles, as the dinosaurs were); not to mention the fact that it would have been functionally useless as a muscle. Evolving this type of novel muscle would have required an unlikely amount of muscle division and reorientation. Granted, this does not mean that these dinosaurs couldn’t have had skin flaps on the sides of their faces to keep food from tumbling out.
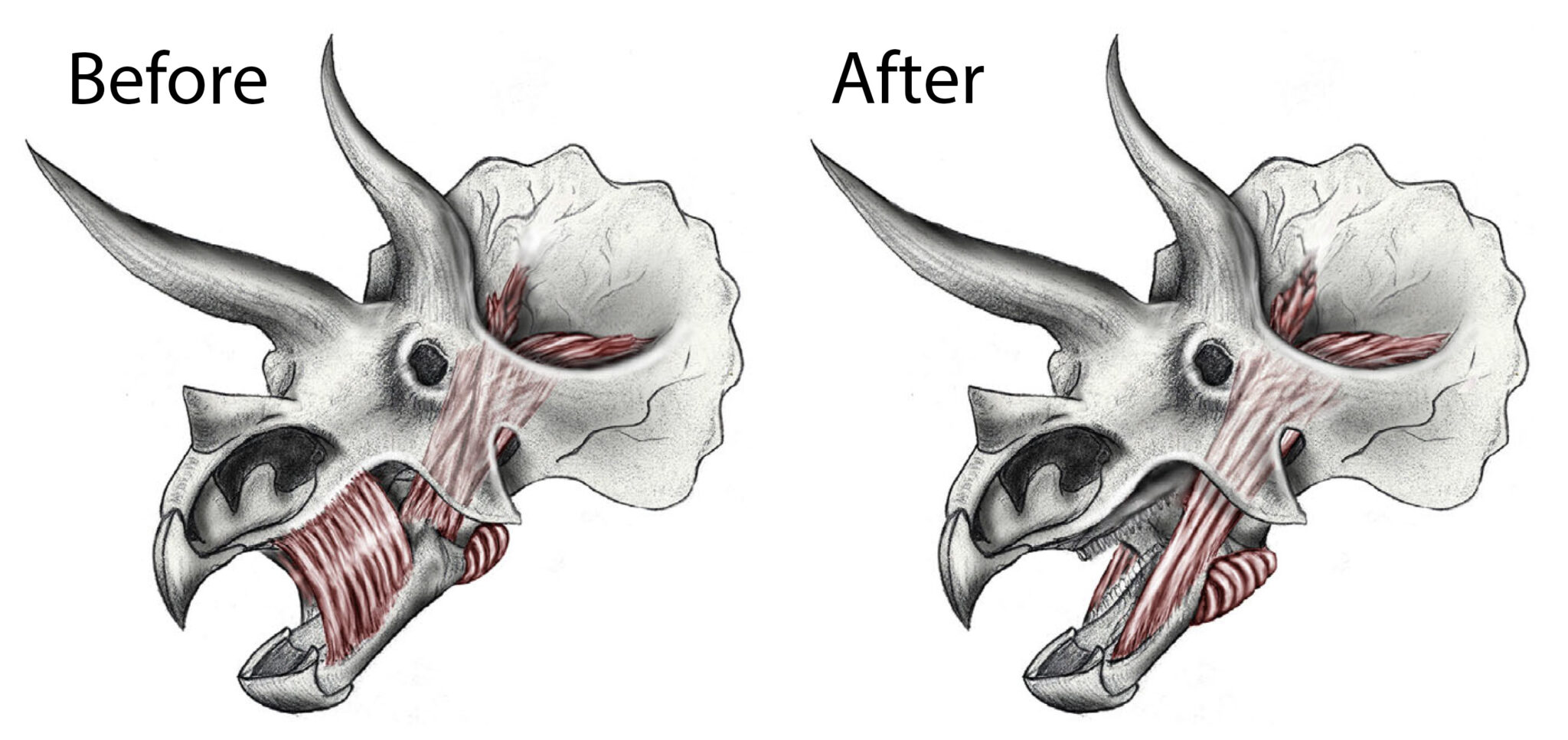
In my paper (Nabavizadeh, 2018), I’ve proposed a new reconstruction of head muscles that takes an already existing layer of muscle on the outer surface of the jaw and extends it forward on the lower jaw as a large fan of muscle (Fig. 2). This muscle fan would attach onto that same ridge that was previously proposed as an attachment of the alleged “cheek” muscle. A large, outwardly-directed jugal (i.e., “cheek bone”) in many ornithischians, especially the horned ceratopsians, was also a dead give-away for the likely presence of a large muscle body to run through and extend forward and down to the lower jaw.
Although at first glance this muscle arrangement seems odd, it is actually seen in a few other extinct animals, particularly those that implement a major palinal (i.e., backward-oriented) feeding stroke in their jaw mechanisms, such as the herbivorous early relatives of mammals known as dicynodonts (King, 1989) and small extinct mammals called multituberculates (Gambaryan and Kielan-Jaworowska, 1995). A fan of muscle was used for palinal feeding in these animals as a support system to lift the entire jaw as one unit to then pull the jaw back while in occlusion. Incidentally, many ornithischians (e.g., the horned ceratopsians, the “duck-billed” hadrosaurs, and the armored ankylosaurs) also incorporated a palinal feeding mechanism themselves, as has been shown by tooth wear analyses (Weishampel, 1984; Varriale, 2011; 2016; Mallon and Anderson, 2014; Osi et al., 2014; Nabavizadeh, 2014; 2016; Nabavizadeh and Weishampel, 2016). Yet, in the original reconstruction, their head muscles were constricted at the back of the jaw, which would not have allowed the jaw to be pulled backward when the animal bit down on something.
This new muscle arrangement would have greatly improved jaw leverage when the animal munched on plants. It also allows for bilateral jaw curling in some of these dinosaurs; specifically, the hadrosaurs and ankylosaurs. With this muscle orientation, the so-called “cheeks” as we know them in large herbivorous mammals were likely not present in ornithischians. Rather, although not its primary function, this large muscle would have secondarily helped keep food in the mouth while it is feeding, as mammal cheeks assist in doing. As a result, this study rewrites how we think of the overall feeding mechanics of these massive herbivores and it also helps us rethink exactly what these animals would have looked like in the flesh—especially because all we have to work with are the bare bones they left behind.
References
Galton PM. 1973. The cheeks of ornithischian dinosaurs. Lethaia 6:67–89.
Holliday CM. 2009. New insights into dinosaur jaw muscle anatomy. Anat Rec 292:1246–1265.
Gambaryan PP, Kielan-Jaworowska Z. 1995. Masticatory musculature of Asian taeniolabidoid multituberculate mammals. Acta Palaeontol Pol 40:1.
King GM, Oelofsen BW, Rubidge BS. 1989. The evolution of the dicynodont feeding system. Zool J Linn Soc 96(2):185–211.
Mallon JC, Anderson JS. 2014. The functional and palaeoecological implications of tooth morphology and wear for the megaherbivorous dinosaurs from the Dinosaur Park Formation (upper Campanian) of Alberta, Canada. PLoS One 9(6):e98605.
Nabavizadeh A. 2014. Hadrosauroid jaw mechanics and the functional significance of the predentary bone. In: Evans D, Eberth D, editors. The hadrosaurs: Proceedings of the International Hadrosaur Symposium. Bloomington: Indiana University Press. p 467–482.
Nabavizadeh A. 2016. Evolutionary trends in the jaw adductor mechanics of ornithischian dinosaurs. Anat Rec 299:271–294.
Nabavizadeh A, Weishampel DB. 2016. The predentary bone and its significance in the evolution of feeding mechanisms in ornithischian dinosaurs. Anat Rec 299(10):1358–1388.
Nabavizadeh, A. 2018. New reconstruction of cranial musculature in ornithischian dinosaurs: implications for feeding mechanisms and buccal anatomy. Anat Rec.
Ösi A, Barrett PM, Földes T, Tokai R. 2014. Wear pattern, dental function, and jaw mechanism in the Late Cretaceous ankylosaur Hungarosaurus. Anat Rec 297:1165–1180.
Varriale FJ. 2011. Dental microwear and the evolution of mastication in ceratopsian dinosaurs. Doctoral dissertation. Baltimore, Maryland: Johns Hopkins University School of Medicine.
Varriale FJ. 2016. Dental microwear reveals mammal-like chewing in the neoceratopsian dinosaur Leptoceratops gracilis. PeerJ 4:e2132.
Weishampel DB. 1984. Evolution of jaw mechanisms in ornithopod dinosaurs. Adv Anat Embr Cell Biol 87:1–109.
Witmer LM. 1995. The extant phylogenetic bracket and the importance of reconstructing soft tissues in fossils. Functional morphology in vertebrate paleontology. Cambridge: Cambridge University Press. Vol. 1. p 19–33.
Featured Image: Comparison of previous head muscle reconstruction (on left, showing hypothetical “cheek” muscle) and new head muscle reconstruction (on right) in the horned ceratopsian dinosaur Triceratops. Modified from Nabavizadeh (2018).
About the Author
Ali Nabavizadeh, PhD, is an assistant professor of anatomy in the Department of Biomedical Sciences at Cooper Medical School of Rowan University. His research investigates comparative anatomy and evolution of herbivorous dinosaurs; specifically, their cranial muscles and the functional morphology of their feeding mechanisms. He also expands these studies to other large herbivorous vertebrates, including dicynodonts and elephants.